
Research
These are some of the areas in which we have worked and continue to have new questions, with answers that continue to surprise. Each of the topics has work underway and plenty of opportunities for making new contributions. Our interests are wide-ranging, however, so if you have something different and exciting in mind, we are happy to discuss possibilities.
Developmental genetics of pigment pattern formation

The vertebrate skin is among the most variable organs in nature, and pigment patterns are among its most striking visible traits. Across ectothermic vertebrates these patterns arise from the spatial arrangement of several classes of neural crest–derived pigment cells. In zebrafish black melanophores and blue-iridescent iridophores make up the dark stripes whereas yellow/orange xanthophores and yellow iridescent iridophores form the light "interstripes." The pigment pattern of zebrafish was a textbook example of vertebrate pattern when our lab began this work, but how this pattern formed was not known. Were stripes specified by pre-existing spatial cues in the surrounding tissue, or did they emerge from active interactions among pigment cells themselves?
Our first contribution came from a mutant with defective stripes — panther — isolated in a forward genetic screen. We showed the gene corresponds to fms (now csf1ra), encoding a receptor tyrosine kinase required for xanthophore development, and that zebrafish lacking xanthophores fail to form proper melanophore stripes. Cell transplants and temperature-sensitive alleles established that xanthophores act non-autonomously and are required throughout development to maintain the striped arrangement of melanophores — zebrafish stripes are not painted onto the body by a pre-existing template but depend instead on active interactions among pigment cells. Subsequent work identified molecular components of these interactions, including adhesion molecules, Delta–Notch signaling, and long-range communication via cellular projections.

wild-type
csf1ra mutant
Interactions among pigment cell classes are an engine to form pattern, amplifying and refining initial biases on when and where the cells develop, a phenomenon with relevance to many types of tissues. Wild-type stripes form when all three classes of pigment cells are present. If even one is missing, stripes cannot form.
Two further lines of work extended this model. First, iridophores — originally thought to play a passive role — turned out to be critical actors. They differentiate first, emerge specifically in the prospective interstripe, themselves express the xanthogenic factor Csf1a, and when ablated disrupt the development of both other chromatophore classes.

Here iridescent iridophores populate the flank in a wild-type zebrafish and contribute to setting the number and orientation of stripes that form. A great advantage to pigmentation is that we can see the behaviours of individual cells and watch phenotypes unfold even in the living animal. Images here are compiled over 30 days in the development of a single individual,
Second, the tissue environment in which pigment cells reside also matters: mutants for the zinc-finger gene basonuclin-2 lack a population of dermal cells and also lack body stripes. Single-cell transcriptomic analysis of zebrafish skin resolved the hypodermis — a thin mesenchymal layer between epidermis and myotome — as a critical permissive tissue, supplying signals that promote pigment cell differentiation, survival, and proliferation, and so allowing patterning interactions to play-out amongst the pigment cells themselves.

Zebrafish skin cell types at single cell resolution and interactions among pigment cells and between pigment cells and their tissue environment, inferred from experimental manipulations to test hypotheses prompted by phenotypes at both whole animal and transcriptomic levels. (Aman et al. 2023)
Current efforts focus on roles for bioelectric signaling and other mechanisms in chromatophore interactions, controls of hypodermal development, and functions of "new" patterning genes found in forward genetic screens and genome-wide association studies.
Research Articles — Parichy et al. Development, 2000; Parichy & Turner Development, 2003; Lang et al. PLoS Genetics, 2009; Eom et al. PLoS Genetics, 2012; Hamada et al. PCMR, 2013; Patterson & Parichy PLoS Genetics, 2013; Eom et al. eLife, 2015; Eom et al. Science, 2017; Eom et al. Dev Biol, 2021; Aman et al. eLife, 2023.
Review Article — Patterson & Parichy Ann Rev Genetics, 2019.
Stem cells, regeneration and cell type diversification
The neural crest is a defining innovation of vertebrates — a transient embryonic population whose derivatives include much of the peripheral nervous system, the craniofacial skeleton, and the pigment cells of the skin. Disruptions in neural crest development and homeostasis are the cause of numerous congenital disorders (like Treacher-Collins syndrome), acquired disease states (Guillain-Barré Syndrome, Charcot-Marie-Tooth Disease, vitiligo and others), and cancers including melanoma.
When the lab began working in this area, neural crest cells were thought to differentiate directly into their derivatives during early embryogenesis. That framing left several questions open. How are the multiple chromatophore classes specified during the larva-to-adult transformation, when most of the adult pattern actually forms? Where do replacement cells come from when adult tissues are damaged in species capable of regeneration? And how do entirely new cell types arise within this lineage over evolutionary time?
A central contribution of our work has been to show that many adult neural-crest derivatives arise not directly from the embryonic neural crest but from latent stem cells established during embryogenesis, maintained in post-embryonic niches, and recruited to differentiate later in development or following injury. We identified peripheral nerves and ganglia as a niche, and showed that precursors first associated with peripheral nerves migrate to the skin during the larva-to-adult transformation, where they give rise to adult melanophores, iridophores and likely neurons and glia — while also contributing cells during regeneration.



Post-embryonic neural crest derived progenitors, moving internally, migrating to the skin to differentiate, and dividing once reaching the hypodermis (Budi et al. 2011).

Post-embryonic neural crest lineages and progenitors in zebrafish (Patterson & Parichy 2019)
Adult xanthophores, however, follow a different route: they derive in large part from embryonic xanthophores that lose their pigment, persist in a cryptic state through larval development, and re-acquire a different type of pigment in the interstripe. The adult pattern is thus built from cells with markedly different developmental histories — some recruited from latent niches, others persisting and reprogrammed in place.

Trajectory analysis of post-embryonic neural crest lineage by scRNA-Seq and lineage analysis of xanthophores in vivo (Saunders et al. 2019; McMenamin et al. 2014).

The alpha tubulin mutant, puma, shows what happens when post-embryonic stem cells are not recruited successfully (Parichy & Turner 2003; Larson et al. 2010).
Three of our more recent directions extend this framework toward cell-type diversification within the adult lineage itself. (i) Thyroid hormone exerts cell type–specific effects, simultaneously promoting xanthophore differentiation and repressing melanophore proliferation, regulating adjacent cell classes in opposing directions through a single global signal. (ii) What had been treated as a single iridophore type in zebrafish in fact comprises distinct crystallotypes — stripe and interstripe iridophores with different reflecting-platelet architectures, optical properties, and transcriptomic states — that arise by differentiation in situ from a common precursor pool. (iii) Entirely new cell types that arise within this lineage: white pigment cells of the zebrafish dorsal fin (melanoleucophores) develop directly from melanophores through a fate conversion driven by intersecting BMP and Agouti signals, with melanin degraded and a guanine-crystal pathway deployed in its place — a striking case of natural transdifferentiation. White cells in a different fin arise instead from yellow xanthophore-like progenitors and depend on a different chemistry — pale and colorless pteridines requiring sepiapterin reductase. Superficially similar leucophores in another species share pteridines but not other features, or use other purines and very different mechanisms. These multiple, developmentally and biochemically distinct ways to make a cell of a given color, scattered across phylogeny, provide a framework for studying cell-type evolution in its own right.
Discovery of new neural crest derived cell types, gene regulatory programs, and evolution by terminal addition in zebrafish (Lewis et al. 2019; Huang et al. 2025; Huang et al. 2026).

Current efforts focus on how gene regulatory programs drive diversification in iridophores and other cell types within ontogenies, how these programs and cells have evolved, stem cell niches and regulatory mechanisms for recruiting cells during regeneration, and how homeostatic mechanisms regulate populations of differentiated cells and prevent overgrowth and cancers.

Super-resolution imaging of pavement-like iridophores within light interstripes and more sparsely arranged iridophores of dark stripes. At right are wild-type, yellowish interstripe iridophores that have developed in a mutant that otherwise replaces these cells with blue stripe-like iridophores.
Research Articles — Parichy et al. Development, 1999; Parichy & Turner Dev Biol, 2003; Parichy et al. Dev Biol, 2003; Budi et al. Development, 2008; Larson et al. Dev Biol, 2010; Budi et al. PLoS Genet, 2011; McMenamin et al. Science, 2014; Saunders et al. eLife, 2019; Gur et al. Nature Comm, 2020; Huang et al. PNAS, 2025; Korzeniwsky et al. PCMR, 2025.
Review — Patterson & Parichy Ann Rev Genetics, 2019.

Progenitor cell at super-resolution (Korzeniwsky et al. 2025).
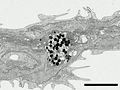
Focused Ion Beam-SEM through a cell transdifferentiating from melanophore to melanoleucophore (Huang et al. 2025).
Evolution of pattern-forming mechanisms and their organismal consequences

Within the zebrafish genus Danio, adult pigment patterns are terrifically diverse — horizontal stripes, vertical bars, spots, mottled patterns, and nearly uniform distributions of pigment cells. When the lab began comparative work, none of this variation had been linked to genes or cell behaviors. Two questions organized our approach: would loci identified in zebrafish predict genes modified during natural pattern diversification? And how do allelic changes translate into cellular outcomes that shape the adult pigment pattern? Early work used interspecific hybrids and targeted genetics to identify molecular pathways, and analyses of cell behaviors in the living fish to uncover changes in morphogenesis, differentiation and stem cell contributions to pattern diversification.
A conceptual advance was that pattern diversification often hinges on timing. In pearl danio D. albolineatus, cis-regulatory variation at csf1a elevates expression of the xanthogenic factor Csf1 and causes xanthophores to differentiate precociously — before iridophores have a chance to develop. Driving Csf1 similarly in zebrafish recapitulated much of the D. albolineatus phenotype. We termed this a “priority effect” — once one chromatophore class differentiates, the interactions it has constrain what follows. A parallel mechanism operates in D. nigrofasciatus, whose attenuated stripes reflect cis-regulatory evolution at Endothelin gene edn3b that curtails iridophore population expansion and for this reason the downstream reiteration of melanophore stripes.


Reduced Edn3b signaling attenuates iridophore development and leads to early cessation of stem cell recruitment to stripe pattern of D. nigrofasciatus (Quigley et al. 2004; Spiewak et al. 2018).
Three more recent extensions integrate this comparative framework with biochemistry, evolutionary genetics, and behavior. (i) Quantitative analyses of Danio "morphospace," combined with QTL crosses between species of divergent pattern, show that pattern variation has a tractable genetic architecture. (ii) Work on carotenoid-based fin coloration in D. albolineatus has characterized the pigments themselves and the enzymes that make them. This includes a vertebrate-wide pathway, shared between fishes and birds, that converts yellow carotenoids to red. A proximodistal BMP signaling gradient then specifies xanthophore or erythrophore fates from a common progenitor at different thresholds. The same logic is deployed in the distantly related Tanichthys albonubes, and acts on different cell types in zebrafish. Connecting mechanism to function, female D. albolineatus prefer to shoal with "super-red" BMP-pathway mutant males — building on earlier work establishing pattern-dependent shoaling in zebrafish. (iii) Collaborative work in anemonefish shows that cellular logic uncovered in Danio recurs across teleosts separated by more than 200 million years of divergence.
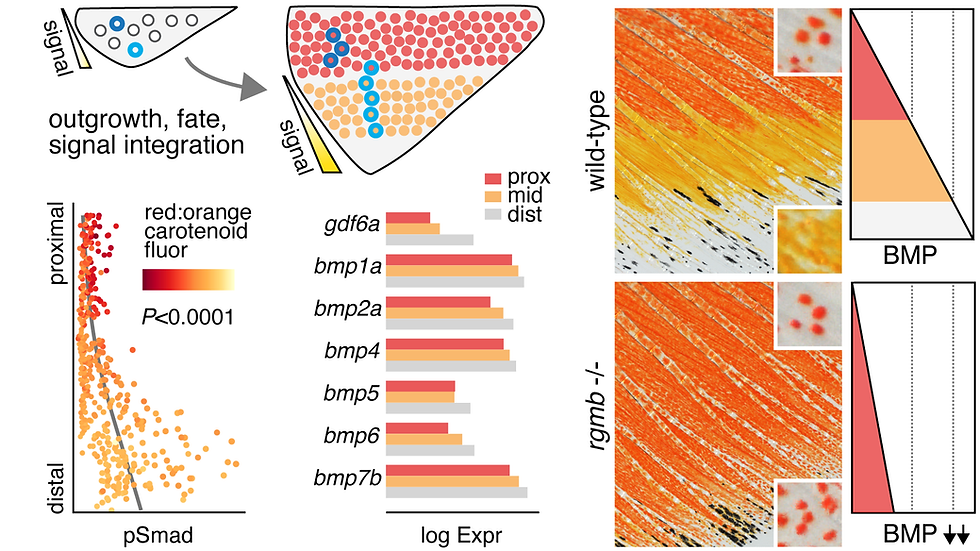
BMP signaling gradient determines erythrophore vs. xanthophore fates and carotenoid phenotypes in pearl danio (Huang et al. 2026).
Current efforts seek to understand how evolutionary changes in pigment cell interactions have contributed to alternative pattern states across species — by analyzing known pathways across species using experimental manipulations and by screening new candidates derived from single cell transcriptomic and other approaches.

Development of a "chain link" pattern from zebrafish-like stripes in D. dangila, which co-occurs with zebrafish in the wild (Engeszer et al. 2007).
We are also using the guppy — with a long history of studies focused on natural and sexual selection — to uncover how positional information interacts with hormonal and other mechanisms to encode genetically variable patterns within natural populations, and how domestication has led to the emergence of variants with remarkable perturbations to the natural state. Finally, additional efforts seek to understand mechanisms of scale ornamentation in minnows as well as entirely unexplored modes of pattern development in marine fishes, especially fairy wrasses.

The remarkable pattern variation of guppy, with just a few wild-types representing a single population, and two out of dozens of "fancy" domesticated forms.
Research articles —
-
Danio: Parichy & Johnson Dev Genes Evol, 2001; Quigley et al. Development, 2004; Quigley et al. Development, 2005; Mills et al. Development, 2007; McMenamin et al. Science, 2014; Patterson et al. Nature Comm, 2014; Spiewak et al. PLoS Genetics, 2018; McCluskey et al. PLoS Genetics, 2021; McCluskey et al, BioOpen, 2021; Huang et al. Curr Biol, 2026
-
zebrafish natural history: Engeszer et al. Zebrafish, 2007
-
behavior: Engeszer et al., Curr Biol 2004; Engeszer et al. PNAS, 2008
-
anemone fish: Salis et al. PNAS 2021, Klann et al. Nature Comm, 2026
Reviews — Parichy Curr Opinion Cell Biol, 2021; Parichy eLife, 2015

Extraordinary patterns of marine wrasses, emblematic of pattern variants found in marine but not freshwater fishes.
Integrated phenotypes: endocrine, paracrine and environmental factors that coordinate adult trait development

Early larva, mid-stage larva and adult zebrafish. Insets show size relative to adult.
Most of vertebrate developmental biology has been a science of embryos. Yet much of what makes an adult organism — the proportions of its skeleton, the architecture of its skin, its size, its appendages, the elaboration of structures laid down only as rudiments during embryogenesis — is built during post-embryonic stages. In fishes these stages span the larva-to-juvenile transformation; across vertebrates they are a frequent target of selection and in humans the source of many congenital and acquired disorders. When the lab began working in this area, no rigorous staging system captured post-embryonic development of zebrafish. We developed a post-embryonic staging series — a "normal table" of development — as a community resource, enabling rigor and reproducibility for work at these stages and serving as a model for analogous tables in other species.

Growth hormone mutant vizzini with a wild-type sibling. Besides its dwarf stature it overgrows adipose tissue (McMenamin et al. 2013)
A consistent finding across multiple traits is that adult form depends on signals integrating growth, differentiation, and homeostasis at a whole-organism scale, often deployed in late-arising contexts. Zebrafish dwarf mutants identified roles for trpm7, encoding a TRP-family ion channel required for skeletogenesis through ion homeostasis rather than classical patterning. The puma mutant — with a missense substitution in alpha tubulin Tuba8l3a — showed that a single cytoskeletal lesion can produce coordinate defects in adult oligodendrocytes, Schwann cells, pigment patterning, and craniofacial morphology, with early larvae forming myelin and glia normally only to develop severe defects during the larva-to-adult transformation — a model for late-arising de- and remyelination phenotypes. Growth hormone signaling, dispensable for embryogenesis, becomes essential at feeding to coordinate body size with adipose dynamics: the GH1 vizzini mutant is severely growth-defective yet attains developmental milestones at smaller-than-typical sizes, with subcutaneous and visceral adipocyte hypertrophy and impaired lipid mobilization.
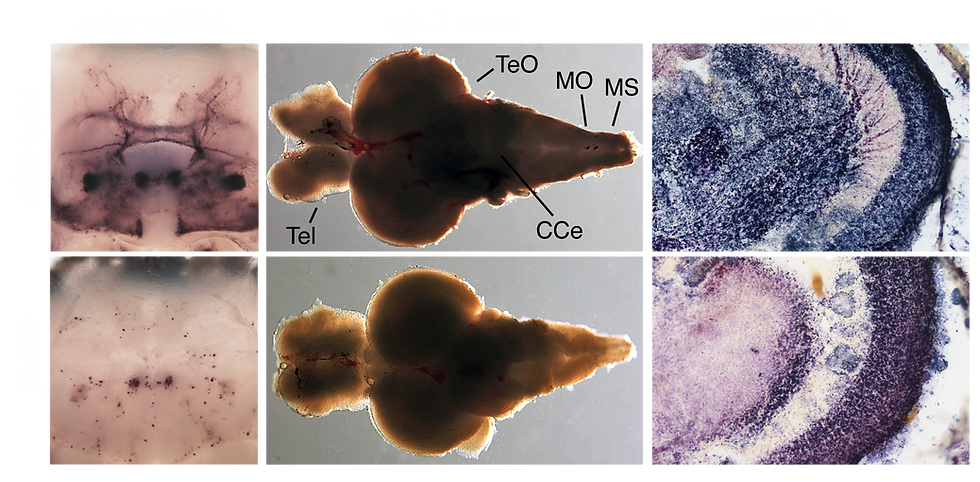
Defective CNS myelination in the tubulin mutant puma accompanies defects in pigmentation and other traits (Larson et al. 2010)
Most extensively studied has been thyroid hormone, which simultaneously promotes xanthophore differentiation and represses melanophore proliferation through a single global signal, with TH dependence itself evolving across species, and a wide variety of additional traits subject to its influence. Among these is scale development, which is dramatically altered in hypothyroid fish both in timing and pattern.

Defective scale pattern formation without thyroid hormone (Aman et al. 2021).
When we started, the cellular composition of post-embryonic teleost skin was poorly resolved, and it was uncertain whether mechanisms patterning fish scales bore deep correspondence with those patterning amniote skin appendages such as feathers and hair. Live imaging through scale ontogeny showed that scales arise sequentially across the flank in an invariant row-and-column progression, with each scale initiated by a Wnt/β-catenin–dependent epithelial signaling center; reciprocal Wnt–Eda interactions, Fgf-dependent differentiation, and a late Hedgehog-dependent program of collective epidermal invagination together account for both scale induction and morphogenesis — placing fish scales squarely within a shared vertebrate program for skin appendage development. Single-cell transcriptomic profiling then resolved skin cell types at high resolution, uncovered a previously undescribed population of basal epidermal cells expressing enamel matrix protein transcripts, and established that thyroid hormone acts on basal epidermal cells to drive Pdgfα-dependent dermal stratification while Eda triggers Fgf20a-mediated scale-forming-cell differentiation.

Thyroid hormone–triggered cascade of interactions underlying scale formation (Aman et al. 2023).
This work exemplifies how trainee-driven curiosity and discovery science can uncover new and interesting biology, made possible by the unusual resources at our disposal. Many exciting questions in this area—of both basic and biomedical relevance—would not have been evident without exploring these later stages of development. Much remains to be done.

Mosaically labeled dermal cells at two display intensities, migrating away from dermal condensates (Aman et al. unpubl).
Additional, newer efforts are aimed at how traits are integrated not just within the animal, but with the environment more broadly, especially through modulation of endocrine systems and life history transitions. For example, we know from our work with zebrafish that development of pigmentation can be partly decoupled from the development of other traits depending on environmental conditions. We are now working to extend these efforts to understand mechanistic underpinnings, using zebrafish as well as other species and other traits. Such insights will be critically important not only for better understanding present-day adult phenotypes and their variation, but also how organisms are likely to respond to changing environments.

Probabilities of pigment pattern stage transitions relative to overall developmental stages (measured by standard length) in zebrafish (Parichy et al. 2009).
Research Articles —
-
zebrafish staging and post-embryonic development: Parichy et al. Dev Dynamics, 2009; McMenamin et al. Methods Cell Biol, 2016
-
trpm7: Elizondo et al. Curr Biol, 2005; Elizondo et al. Endocrinology, 2010
-
growth hormone and adiposity: McMenamin et al. Endocrinology, 2013
-
TH, scales: McMenamin et al. Science, 2014; Aman et al. eLife, 2019; Aman et al. Dev Biol, 2021; Aman et al. eLife, 2023
Population biology and genetics of danios

Field work on zebrafish in West Bengal.
The zebrafish became a biomedical model not because of what was known about its ecology and behavior in nature, but in spite of how little was known. Like most laboratory model organisms, it was chosen for traits convenient to laboratory science — easy to breed, transparent embryo, short generation time, large clutches — and was for decades studied largely without reference to the species' natural setting. Yet a model organism is also a species with a natural history, and three considerations make that history matter for the kind of work this lab does.
First, the form, behavior, and genetic variation of zebrafish in the laboratory reflect a long history of selection in particular environments, and a deeper understanding of those environments helps to interpret what we see in the tank. Second, comparison across populations, across closely related species, and across the broader phylogeny of Danio is one of the strongest tools available for asking which features of zebrafish biology are general and which are derived. Third, the patterns of allelic variation that segregate in nature are not the same as those recovered by laboratory mutagenesis, and understanding the genetic architecture of naturally occurring traits is essential for translating between lab and field, and between zebrafish and other vertebrates. When the lab began working in this area, the field had only a sparse and outdated literature on wild zebrafish, an unclear picture of its habitat and life cycle, and almost no information on the genetic basis of variation in pigmentation or other traits across natural populations or closely related species.


A shoal of zebrafish in its preferred habitat. Meghalaya, India. (Engeszer et al. 2007)
Our first contribution to this area was a field survey of zebrafish habitat and natural history across northeastern India. Across sites in West Bengal, Assam, Meghalaya, and Orissa, ranging from sea level to nearly 1800 m elevation, we found zebrafish reliably associated with slow or still water — silt-bottomed pools at stream margins, swampy areas, and flooded rice paddies — and across remarkable ranges of temperature (24.6–38.6 °C) and pH (5.9–8.1). We documented co-occurring fish fauna in detail, identifying likely competitors (other small cyprinids in the genera Danio, Devario, Esomus, and Puntius) and predators (snakeheads, knifefish, catfish, needlefish, and aquatic dragonfly larvae). We proposed a life-cycle model in which adults occupy streams between monsoons, move into seasonal flooded habitats to breed, and larvae and juveniles develop in these ephemeral nursery pools before returning to the streams.


Catching fish.
We also documented the rapid and ongoing degradation of zebrafish habitat through mining runoff, detergent contamination, and fishing with industrial poisons — a sobering observation for a species often described as hardy. A subsequent synthesis extended this picture, situating zebrafish within the considerable genetic diversity of natural populations (lab strains capture only a small fraction of total variation), and the broader phylogeny of subfamily Danioninae, in which Danio rerio is one of roughly twenty species spanning an unusual range of body sizes — from miniaturized Danionella to D. dangila several times larger than zebrafish — and a striking diversity of pigment patterns. This comparative framing has anchored much of the lab's later work on Danio diversity.

Danio patterns and phylogeny (Parichy 2015). D. aff. kyathit is now D. quagga.
Two more recent studies bring genetic analysis to bear on questions raised by this comparative framework. The first concerns sister taxa — Danio quagga and D. kyathit, probably a single biological species — that differ dramatically in adult pigmentation: D. quagga develops horizontal stripes much like zebrafish, while D. kyathit develops a spotted pattern. Strikingly, the D. kyathit phenotype occupies a region of pigment-pattern morphospace also occupied by several single-locus zebrafish mutants, which might suggest that the species difference also resolves to a single major locus. Reduced-representation sequencing of F2 hybrids told a different story. Pattern variation between the taxa — both at the level of overall morphology and at the level of cellular phenotypes — maps to multiple quantitative trait loci, and abundant segregating variation persists within each taxon as well. The genetic architecture of the difference is therefore more complex than that of superficially similar laboratory mutants, illustrating the gap between simple loss-of-function alleles and the alleles that actually contribute to natural divergence. A parallel result emerged from work on D. albolineatus fin pattern. The recurring lesson is that allelic variation in nature is architecturally more complex than the mutant alleles isolated in laboratory genetic screens.

D. quagga (stripes) and D. kyathit (spots).
Current efforts focus on naturally occurring allelic variation for pigment pattern between inbred strains of zebrafish, by genome-wide association using new metrics and tools for pattern analysis. Here, too, an oligogenic basis of variation is apparent and candidate gene testing by CRISPR screening has already identified candidate genes within critical intervals that have pigmentary phenotypes, despite these genes not being associated with pigmentation previously. Through efforts like these we aim to gain a fuller appreciation of the causes and consequences of population level variability.
Research Articles — Engeszer et al. Zebrafish, 2007: McCluskey et al. PLoS Genetics, 2021; Huang et al. Curr Biol, 2026; Lu et al. bioRxiv, 2026.
Review — Parichy eLife, 2015
